Introduction

As the sister group of animals and part of the eukaryotic crown group that radiated about a billion years ago, the fungi constitute an independent group equal in rank to that of plants and animals. They share with animals the ability to export hydrolytic enzymes that break down biopolymers, which can be absorbed for nutrition. Rather than requiring a stomach to accomplish digestion, fungi live in their own food supply and simply grow into new food as the local environment becomes nutrient depleted.
Most biologists have seen dense filamentous fungal colonies growing on rich nutrient agar plates, but in nature the filaments can be much longer and the colonies less dense. When one of the filaments contacts a food supply, the entire colony mobilizes and reallocates resources to exploit the new food. Should all food become depleted, sporulation is triggered. Although the fungal filaments and spores are microscopic, the colony can be very large with individuals of some species rivaling the mass of the largest animals or plants.

Click on an image to view larger version & data in a new window
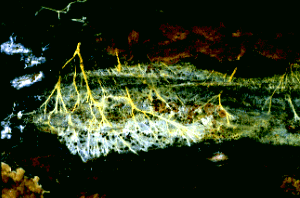
Figure 1: Hyphae of a wood-decaying fungus found growing on the underside of a fallen log. The metabolically active hyphae have secreted droplets on their surfaces. Copyright © M. Blackwell 1996.
Prior to mating in sexual reproduction, individual fungi communicate with other individuals chemically via pheromones. In every phylum at least one pheromone has been characterized, and they range from sesquiterpines and derivatives of the carotenoid pathway in chytridiomycetes and zygomycetes to oligopeptides in ascomycetes and basidiomycetes.Within their varied natural habitats fungi usually are the primary decomposer organisms present. Many species are free-living saprobes (users of carbon fixed by other organisms) in woody substrates, soils, leaf litter, dead animals, and animal exudates. The large cavities eaten out of living trees by wood-decaying fungi provide nest holes for a variety of animals, and extinction of the ivory billed woodpecker was due in large part to loss, through human activity, of nesting trees in bottom land hardwoods. In some low nitrogen environments several independent groups of fungi have adaptations such as nooses and sticky knobs with which to trap and degrade nematodes and other small animals. A number of references on fungal ecology are available (Carroll and Wicklow, 1992; Cooke and Whipps, 1993; Dix and Webster, 1995).
However, many other fungi are biotrophs, and in this role a number of successful groups form symbiotic associations with plants (including algae), animals (especially arthropods), and prokaryotes. Examples are lichens, mycorrhizae, and leaf and stem endophytes. Although lichens may seem infrequent in polluted cities, they can form the dominant vegetation in nordic environments, and there is a better than 80% chance that any plant you find is mycorrhizal. Leaf and stem endophytes are a more recent discovery, and some of these fungi can protect the plants they inhabit from herbivory and even influence flowering and other aspects of plant reproductive biology. Fungi are our most important plant pathogens, and include rusts, smuts, and many ascomycetes such as the agents of Dutch elm disease and chestnut blight. Among the other well known associations are fungal parasites of animals. Humans, for example, may succumb to diseases caused by Pneumocystis (a type of pneumonia that affects individuals with supressed immune systems), Coccidioides (valley fever), Ajellomyces (blastomycosis and histoplasmosis), and Cryptococcus (cryptococcosis) (Kwon-Chung and Bennett, 1992).
Click on an image to view larger version & data in a new window
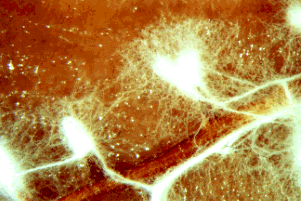
Figure 2: The fluffy white hyphae of the mycorrhizal fungus Rhizopogon rubescens has enveloped the smaller roots of a Virginia pine seedling. Note that some of the mycelium extends out into the surrounding environment. Copyright © J. B. Anderson 1996.
Click on an image to view larger version & data in a new window

Figure 3: Entomophthora, "destroyer of insects", is the agent of a fungual infection that kills flies. After their death the fungal growth erupts through the fly cuticle, and dispersal by forcible spore discharge is a source of inoculum for infection of new flies. Copyright © G. L. Barron 1996.
Fungal spores may be actively or passively released for dispersal by several effective methods. The air we breathe is filled with spores of species that are air dispersed. These usually are species that produce large numbers of spores, and examples include many species pathogenic on agricultural crops and trees. Other species are adapted for dispersal within or on the surfaces of animals (particularly arthropods). Some fungi are rain splash or flowing water dispersed. In a few cases the forcible release of spores is sufficient to serve as the dispersal method as well. The function of some spores is not primarily for dispersal, but to allow the organisms to survive as resistant cells during periods when the conditions of the environment are not conducive to growth.Fungi are vital for their ecosystem functions, some of which we have reviewed in the previous paragraphs. In addition a number of fungi are used in the processing and flavoring of foods (baker's and brewer's yeasts, Penicillia in cheese-making) and in production of antibiotics and organic acids. Other fungi produce secondary metabolites such as aflatoxins that may be potent toxins and carcinogens in food of birds, fish, humans, and other mammals.
A few species are studied as model organisms that can be used to gain knowledge of basic processes such as genetics, physiology, biochemistry, and molecular biology with results that are applicable to many organisms (Taylor et al., 1993). Some of the fungi that have been intensively studied in this way include Saccharomyces cereviseae, Neurospora crassa, and Ustilago maydis.
Most phyla appear to be terrestrial in origin, although all major groups have invaded marine and freshwater habitats. An exception to this generality is the flagellum-bearing phyla Chytridiomycota, Blastocladiomycota, and Neocallimastigomycota (collectively referred to as chytrids), which probably had an aquatic origin. Extant chytrid species also occur in terrestrial environments as plant pathogenic fungi, soil fungi, and even as anaerobic inhabitants of the guts of herbivores such as cows (all Neocallimastigomycota).
Characteristics

Click on an image to view larger version & data in a new window
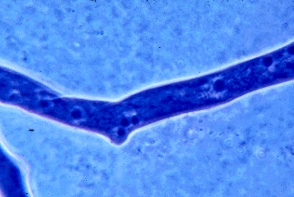
Figure 4: Portion of a hypha of a zygomycete stained with a blue dye to show the many nuclei present. Many other fungi have septations that devide the hyphae into compartments that usually contain one to several nuclei per compartment. Copyright © M. Blackwell 1996.
Click on an image to view larger version & data in a new window
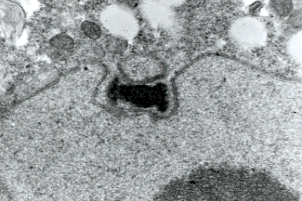
Figure 5: Transmission electron micrograph showing duplicated spindle pole body of a prophase I meiotic nucleus of a basidiomycete Exobasidium. Only chytrids among fungi have centrioles and lack spindle pole bodies. Copyright © Beth Richardson 1996.
Exceptions to this characterization of fungi are well known, and include the following: Most species of chytrids have cells with a single, smooth, posteriorly inserted flagellum at some stage in the life cycle, and centrioles are associated with nuclear division. The life cycles of most chytrids are poorly studied, but some (Blastocladiomycota) are known to have zygotic meiosis (therefore, alternation between haploid and diploid generations). Certain members of Mucoromycotina, Ascomycota, and Basidiomycota may lack hyphal growth during part or all of their life cycles, and, instead, produce budding yeast cells. Most fungal species with yeast growth forms contain only minute amounts of chitin in the walls of the yeast cells. A few species of Ascomycota (Ophiostomataceae) have cellulose in their walls, and certain members of Blastocladiomycota and Entomophthoromycotina lack walls during part of their life cycle (Alexopoulos et al., 1996).Fossil Record

A first attempt to match molecular data on fungal phylogeny to the geological record shows general agreement, but does point out some conflicts between the two types of data (Berbee and Taylor 1993).